在未受到刺激时,神经纤维处于静息状态,此时细胞膜外电位比国内高,细胞膜两侧的电位表现为内副外症,称为静息电位。神经元静息电位与细胞外的大离子浓度和细胞内的假离子浓度有关,而电化学梯度也正是由大甲泵来建立和维持的。 当神经纤维某一部位受到刺激时,细胞膜对纳离子的通透性增强,纳离子内流这个部位的膜两侧出现暂时性的点位变化,表现为内正外负的兴奋状态,此时的膜点位称为动作点位, 而临近的未兴奋部位仍然是内副外正。在兴奋部位和未兴奋部位之间由于电位差的存在而发生电和移动,这样就形成了局部电流。这种局部电流又刺激相近的未兴奋部位发生同样的电位变化,如此进行下去,将兴奋向前传导,后方又恢复为 吸电位。同时需要注意的是,刺激神经纤维上任意一点兴奋会沿着神经纤维向两侧传道,因为刺激位置兴奋后,无论是兴奋位置的左侧还是右侧, 均与兴奋位置存在电位差,均可形成局部电流,将兴奋向外传递,在短暂的延迟后,假离子通道开放,假离子外流又恢复为静息电位。

粉丝22.2万获赞272.6万
相关视频
 05:24查看AI文稿AI文稿
05:24查看AI文稿AI文稿大家好,今天我们学习神经冲动的产生和传导,兴奋在神经纤维上的传导, 大家请看图。一、传导形式据科学家实验,在挖的坐骨神经上,神经纤维膜外放置两个微电极,并将他们连接到同一个电流表上。静息时,电流表指针不偏转,说明神经表面各处电位相等。 当在神经的左侧一端给予四级时,当靠当靠近四级一端先变为负点位,另一侧还是正点位。 两处电位形成了电视差,形成了局部电流,电流流向电位低的一方,电流表指针向左片段。紧接着兴奋沿着神经纤维传导另一侧电极处 变为负点位,而靠近四级一侧恢复为正点位,电流表指针向右偏转,最终两侧点位恢复一致,电流表不再偏转。该实验说明,身份的产生需要一定的刺激,也就是说达到预值。 身份在神经纤维中是以电信号的形式传导的,我们把这种电信号也叫做神经冲动。第二个,静息电位和动作电位及其成因 近期电位内负外正形成原因是假离子外流,神经细胞膜上的假离子通道打开,那离子通道关闭,假离子以协助扩散的方式大量外流,使膜外阳离子浓度高于膜内动作电位 内政外附形成的原因是,当受到刺激时,神经细胞抹上的纳离子通道打开,假离子通道关闭。 纳尼斯以协助扩散的方式大量内流,造成模内阳逆子浓度高于膜外,该部位发生模内外钻石中的变味变化,即由内负外症变为内政外负的兴奋状态。 传导过程在身份部位和胃身份部位的膜内膜内外临近,临近部分之间由于电位差的存在而发生电合的移动,也就是说由正离死变为负离死,这样就产生了局部电流。 止步电流的方向是由途中箭头的方向,止步电流又作为新的刺激,使相近的胃形成部位发生同样的变味变化。如此进行,身份就可以向前传导,后方又因为假离子外流恢复为近期状态。为了使神经元内外离子 浓度平衡,加纳离子泵会吸纳吸甲排纳,主动运输。局部电流的方向在某外由未兴奋部位流向兴奋部位。在某内由兴奋部位流向未兴奋部位,从而形成了局部电流的回路。 兴奋的传导方向由兴奋部位到微兴奋部位,与模内屏幕电流方向一致,在离体神经纤维上双向传导,在反射弧中单向传导。兴奋来自于感受器。 三、在神经纤维上电流表的偏转分析一、刺激 a 点 b 点前先先兴奋, b 点后兴奋,电流表发生两次方向相反的偏转,先左后右。刺激 c 点 bc 等于 cdb 点和地点,同时兴奋电流计不偏转。三、刺激 cd 中点, 地点前胸先兴奋,必点后兴奋,电流表发生两次方向相反的片段,也就是说先右后左。 根据以上的内容的了解,我们来做一个小题,如图,甲是测量神经纤维膜内外的膜内外垫位的装置图以是测的膜垫位的变化曲线。下列相关数数正确的是。我们看一下这个图,这个图中甲图是有甲,图中的 a 是 外政内负,也就是说是假,因为假离子外流,呃那里是通道闭合而导致而导致的间隙变味,也就是土以中的 c 处。 假假图中的 b 是外负内正是因为那里是大量内流而导致成的动作点位。也就是说,假图中的 a 处外负外正内负,是由 于甲离子大量外外流,纳离子、纳离子通到 b 和而导致成的正气变味。是土以中的 a 处,而甲子中的 b 是由于呃外是外副内症是由于纳离子通到打开纳离子大量内流,假离子通到闭合 而造就成了洞子变位,也就是图以中的 c 处。而甲图中 a 到 b 的过程是图以中的 b 处 a, 图甲中 b 所处所 c 处值是。这些点位相当于图以中的 a 点的点位,是错误的,应该是 b 处是洞处点位相当于图以中的 c 点的点位。 b。 若细胞外纳底子浓度适当,身高在适宜条件下刺激土以中 c 点上移,这个是正确的。 c 神经纤维的状态由 a 转变为 b 的过程, 我对假离子的通透性增大是错误的,应该是大离子的通透性增大的。又要画出土、乙的动作定位,需要测量神经三位上多个位点的定位变化,这个是错误的,应该是同一个位点的不同变化。所以这个题应该选的是比 我讲完了,谢谢大家。
43老段01 15:00查看AI文稿AI文稿
15:00查看AI文稿AI文稿hello, 大家好,我是绵绵老师,今天我们来进行摩电位机理和曲线分析的知识讲解。 首先呢,我们要来补充一点小知识,这是细胞膜,细胞外分布的阳离子主要是钠离子。 对于钠离子来说,膜外的浓度要高于膜内,而细胞内分布的阳离子主要是钾粒子。 对于钾离子来说,膜内的浓度要高于膜外,而细胞外分布的阴离子主要是氯离子。细胞内分布的阴离子主要是有机阴离子, 我们用 a 负来表示,比如说一些带负电荷的蛋白质分子,在这个图片当中,我们是用字母 母的大小来代表数目的多少。那这节课呢,我们会涉及到钠离子、假离子的运输,氯离子和 if, 不多加讲解。 第二个要补充的小知识是,我们需要认识到细胞膜上的这四类转用蛋白。第一个是纳离子电压门控通道,正如其名,这是一个转用纳离子的通道蛋白, 他的开放与关闭受到摩电位的调控。第二个是假离子电压门控通道,这是一个转用假离子的通道蛋白, 它的开放与关闭同样受到摩电位的调控。第三个是纳甲泵,它一直处于开放的状态, 每消耗一个 atp 分子呢,它可以向包外泵出三个钠离子,向包内泵入两个钾离子。从钠离子和钾离子的分布情况我们不难看出,它转用钠离子和钾离子都是从低浓度到高浓度, 并且需要消耗能量,所以钠钾泵转用钠离子和钾离子的方式是主动运输。 钠钾苯的作用在于维持膜两侧的离子浓度差,也就是维持包外钠离子浓度高,包内钾离子浓度高的状态。 第四个是非门控假离子通道,这个通道蛋白一直处于开放的状态, 那在安静的状态下呢?纳离子电压门控通道,假离子电压门控通道处于关闭的状态,纳钾泵和非门控假离子通道处于开放的状态。在安静的时候,膜主要对假离子有通透性。 那通过刚刚的分析呢,我们也知道,在细胞内钾离子的浓度要高, 而在细胞外钾离子的浓度要低,这样的话,钾离子就会在浓度差的驱动下通过非门控钾离子通道外流。 外流会使得膜外的阳离子浓度要高于膜内,这样子细胞膜两侧的电位表现 就表现为外正内负。我们把这个定位差叫做进气定位。 简单的小结一下,近吸电位产生的原因在于钾离子向膜外跨膜转,用跨膜用书的方式是协助扩散。没错,第一个是因为它是顺浓度剃度用书,第二个是因因为它需要 非门控假离子通道这个通道蛋白的帮助。现在呢,我给神经纤维某一个部分适宜的刺激,那纳离子电压门控通道就会从关闭的状态变成开放的状态, 假离子电压门控通道依旧处于关闭的状态,那纳离子电压门控通道开放,它会引发纳离子内流。 这个过程呢,首先是少量兴奋性较高的那通道开放,引起少量的纳离子内流,纳离子内流会使得 膜内的负值减小,那膜电位减小到一定的数值值的时候,也就是遇电位的时候,大量的纳通道同时开放,引起大量的内里纳离子内流, 钠离子内流的最终结果会使得膜内的阳离子浓度要高于膜外, 也就是说细胞膜两侧的电位表现为内正外负, 我们把它称作为动作定位。动作定位产生的原因在于纳离子向膜内跨膜转用,跨膜转用的方式依旧是协助扩散, 那接下来呢?这个地方的抹电位,他会向近期电位恢复,所以我们来看一下近期电位恢复的机制。 纳离子电压门控通道处于关闭的状态,而假离子电压门控通道处于开放的状态,那从这里我们可以知道,近期电位恢复的基础就是假离子通过假离子电压门控 通道向膜外流动,那也就是说钾离子向膜外跨膜转用跨膜运输的方式依旧是协助扩散。 那现在呢?我们是已经梳理了近期电位动作定位、近期电位恢复的机制。那在真实的历史上,近期电位和动作定位是怎么被测量出来的?我们来看一下这个图, 在这个图片当中,你能找到两个电极参考,电极置于细胞外,并通过细胞外液接地,使得膜外的电位固定为零毫伏,而记录电极则插入到细胞内,那这样的话,我们就可以通过试播 记录到一个电位差。我们来想一下,在安静的状态下,这个电位差记录到的是一个正值还是一个负值? 在安静的状态下,外正内负, 那我们把细胞外的电位固定为零,那细胞内的电位肯定就更是一个负值,那所以记录到的电位差是一个负值, 那这个细胞它的精细电位呢是负七十毫伏,我们同样呢展示了一些其他细胞的精细电位,我们来 讨论一下复值后面的那个数字代表着什么?它其实代表了细胞膜两侧的电位差,这个数字如果越大,那膜两侧的电位差也就越大,我们就说近习点位越大。 那之后我们来比较近期电位的大小和近期电位的变化,都说的是符号后面那个数字的大小。 好,接着呢,我给这个神经纤维某一个部分一个刺激, 那将会引发纳离子内流,膜内外的电位差就逐渐的缩小至零,并出现反转,反转的结果会 使得膜内的电位要高于膜外,我们把它称作动作电位。那根据记录到的数据,我们可以来画出一条曲线,这条曲线的横坐标是时间,纵坐标是摩电位。 从这个图片当中我们不难看出,这个神经细胞的近期电位是负七十毫伏。近期电位产生的原因在于钾离子通过协助扩散外流, 现在给它施加一个刺激,会使得纳离子电压门控通道开放纳离子内流。哎,这个用处的方式依旧是协助扩散,那纳离子内流会使得细胞 内复制减小。我们把细胞内复制减小的这个过程叫做区级化,也就是说这一段是区级化,那细胞膜区级化之后,他会到达一个动作定位的分值, 也就是这个曲线的最高点,这是动作定位的分值,那接下来这个地方的抹定位将会向近期定位恢复。我们把这个过程叫做负极化, 负极化的机制在于假离子电压门控通道开放,假离子外流用书的方式依旧是协助扩散,那 从这张曲线当中我们也可以看出,它并不是一下子就回到了近习电位,而是需要通过一个阶段,在这个阶段当中, 细胞膜的膜电位,它的负号后面的那个数字要比近系电位更大,那我们在生理学对于此阶段的说明的话是纳甲泵活动增强。 我们来看一下纳钾泵的图解,消耗一个 atp 分子向包外泵出三个钠离子,向包内泵入两个钾离子,也就是相当于把一个近的正电荷泵泵出了包外, 那这样的话就会强化外震内复的一个状态,也就导致了超级化细胞内复制增大的过程叫做超级化。 那后来呢?他会通过一些其他电流,什么电位到达近吸电位, 那在这个图片当中,棕色的部分大家一定要熟知,而绿色的部分你可以简单的了解一下, 那在这里我们需要强调一点,跨膜用书的离子量很少很少,只需要百万分之一的假离子外流就可以使抹电位 到达近期电位。那所以说对于假离子来说,永远都是膜内要高于膜外, 而对于那里子来说,永远都是膜外要高于膜内。 好,分析完了这条曲线,我们接下来说一下动作定位的特点。 动作定位有一个特点,全或无现象,他是说什么的呢?他指的是要使细胞产生动作定位所给的刺激必须达到一定的强度,如果这个刺激没有达到这个强度的临界值,动作定位就不会产生,这说的是 五,那当刺激达到一定的强度时,所产生的动作定位的幅度便到达该细胞动作定位的最大值,不会随着刺激强度的继续增大而增大,这说的是全, 那怎么理解这句话,我们来看一下,这是因为呢?刺激强度只决定摩电位是否能到达预电位。 好,接下来我们根据刚刚的摩垫位曲线图来说明一下,这个神经细胞对应的预电位是负五十五毫伏, 预电位指的是能触发动作电位的膜电位临界值,如果这个刺激可以使细胞膜发生趋激化到预电位,那后续的这些过程 就和刺激强度没有关系。那如果这个刺激无法使细胞膜发生去激化 到预电位,比如说直使细胞膜发生区级化到负六十毫伏, 那这时候呢?细胞膜发生的是轻微的去挤化,这个轻微的去挤化会很快被加强的假离子外流所抵消掉, 那紫色的部分就不会发生,也就是说不会爆发动作定位,这说的就是无。 那我们把,嗯,刚好能使细胞膜发生去挤化到遇电位的刺激叫做遇刺激。通过刚刚的分析我们就知道,能使细胞产生动作电位的有效刺激指的是遇刺激和强度 要大于遇刺激的遇上刺激。 那说了这么多,我们接下来思考两个小问题,第一个问题,动作定位的分值与什么有关?第二个问题,近期定位的大小与什么有关? 那我们来想一下,动作电位产生的原因在于纳离子通过纳离子电压门控通道 内流,那所以动作电位的分子肯定与那通道有关,那纳离子内流它的驱动力是浓度差,所以它也与细胞膜两侧纳离子浓度差有关。 那近吸电位的大小与什么有关呢?近吸电位的原因在于钾离子外流,钾离。
543棉棉老师的生物课堂 13:36查看AI文稿AI文稿
13:36查看AI文稿AI文稿同学们好,接下来我们看第三节有关神经冲动的产生和传导,这讲应该是整个这章的一个重点, 我们看一下主要涉及到哪些方面的内容。两个基本的字典。第一,兴奋他在神经纤维上是怎么产生可传导的? 第二,由于在反射活动完成的过程当中,他需要跨越不同的细胞,因此这样的话,在细胞与细胞之间这种信号又是如何进行传导的?所以这样的话就有两个基本的问题,一是兴奋在神经纤维上的传导,第二个是兴奋在神经 圆之间的一个传递,那么当然还会涉及到一些相应的应用。好,接下来我们 看第一个部分的内容,先看一看兴奋他在神经纤维上的传导应该是怎么完成的。在没有受到刺激的情况之下, 细胞膜内外之间是存在着一个定位的差在里边的,我们知道他应该是一个外增内负,那么这个之间的差值大概应该是在这边负七十的左右,这个时候我们把它叫做一个静息的一个状态, 由于他存在着电位的差,所以我们也把它称之为即化的状态,然后当给予一个刺激之后,这种即化状态就会消失,所以把它叫做去洁化, 不仅仅是消失了,而且他的定位是给反过来了,一直到了正的三十左右吗?因此这样的话他就给反过来,那么原来是一个外震内 的,现在就变成是外负内重的这么一个基本关系。第三,当完成这个兴奋产生之后,后边这个兴奋又恢复到原来的一个状态, 所以这样的话,我们说第三个阶段就是一个负极化的一个过程,所以你看原来是个极化的状态,去极化,然后再负极化,那么就恢复到原来受到刺激之前的这样一种极化的状态。至于这个图上还有个超级化的这个阶段,我们给他 忽略了,所以我们现在主要就是关注这样三个相应的阶段。那么首先这个近期的状态,他为什么会处在一个极化的状态呢? 他跟细胞里边和外边的离子是密切相关的,在细胞里边的阳离子比较多的是假离子, 而因离子主要是蛋白质等等大分子的这个部分在细胞的外边主要应该是钠离子, 然后因离子的话主要是绿离子。那么可能有同学会觉得,为什么细胞外边钠离子的浓度是比较高的,而在细胞的里边是假离子浓度比较高,而不会给换过来呢? 那最主要的是细胞膜上面有一种结构叫做纳甲泵,这种纳甲泵的工作,他将能够保证细胞在外边钠离子浓度高,而在细胞里边钾离子浓度是比较高的。 换过来说,如果细胞里边纳离子浓度高了,纳点笨就会启动工作,工作之后的结果那就是把纳离子给他送到细胞外边去,而同时把钾离子给换到细胞里边来,这样 就保证了两者之间的一个即兴的一个状态,这样也就保证了细胞外边的钠离子浓度是高的,而细胞里边的假离子浓度是比较高的。 呃,在没有受到刺激的情况之下,有一个通道是打开的,这是假离子的通道,所以假离子他是可以顺着浓度往外边进行流的, 那么在假离子外流之后,外边的假离子逐渐会增高,所以他会变成是一个带正电的外震内负,主要就是由于假离子外流的结果。可是假离子外流为什么不会维持一个平衡呢? 那是因为当外边带了正电之后,这个时候假离子再要出去的话,他就有了阻力,细胞里边的负电也将会吸引着他, 所以这个时候的话,一方面是细胞里边钾离子浓度高出去的力量,一方面是外边已经带正点阻止他出去的力量,两者之间达到平衡的状态的时候,那么就处在一个静息的一个状态,这个时候就维持在一个大约 负七十的这样一个相关的状态,那么接下来的话去计划是受到了刺激之后所发生的。为什么会改变电核的分路变成是外负内重呢? 这个时候主要是一个一种离子的作用,那就是纳离子,纳离子内流了,因为细胞外边的纳离子大量的流入到细胞里边来,所以这样的话就改变了原来的外震内腹的状态,而变成是外边是,所以腹的里边 变成是正的这样一个状态,那在恢复的过程当中,甲离子的通道又会打开, 所以这个时候假离子会流出去,那么这样的话大家就会觉得原本应该是属于细胞外边是钠离子浓度高的,细胞里边是假离子浓度高的,可是在聚集化的过程当中,钠离子流到细胞里边来了, 而在负极化的时候,钾离子又流出去了,那么这样的话就会改变细胞内外的纳钾离子的一个浓度, 这个时候谁就开始工作了,那家笨,那家笨开始工作的话,他将能够把这两种离子再给他换回来,仍然换到里边是假离子浓度高,而外边是属于纳离子浓度高的这样一个相关的状态,所以这就是一个兴奋 产生和恢复的一个基本的过程。好,那么一旦这个兴奋产生之后,他向着旁侧进行移动,那么我们看一下, 这个地方是兴奋了,变成是外腹内震,而周边仍然是属于外震内腹的状态,所以我们就可以看到这有正腹,细胞膜里边也有正腹,那就形成了局部的电流,所以电流就应该是会从正的流向腹的这一侧, 所以这样的话局部电流形成,那么就会引起,呃,原来没有兴奋的地方,也就会发生相应的兴奋,那么这样兴奋就开始发生移动。在这的话,有一个地方我们简单的交代一下,呃,如果当这个动作定位传到这个位置的时候,可能 没有同学会觉得两侧都是属于带正的,那么岂不意味着两侧都会形成一个局部的电流?为什么在这边图上只表示了一侧呢? 那是因为刚刚兴奋的部分是不会立刻再进行兴奋的,他有一个不能对自己做出反应的时期,也正因为如此,所以决定了这个兴奋只能什么朝着一个方向进行移动,而不会在两边倒来倒去的倒 好。所以这个是我们所涉及到这个兴奋传导的过程当中。呃,大家应该要注意的一个问题, 但是有一点,如果我们是在某一个中间的部分开始给予的刺激的话,由于两侧的部分都没有兴奋过,因此在这种情况之下,兴奋是能够朝着两个方向进行扩散的。我们 只是说刚兴奋的部位在重新接受到刺激的情况之下,不会立刻进行兴奋,所以这个呢,应该要能够注意到好,这是兴奋的一个产生可传导的问题, 那么也正因为如此,所以我们说在神经纤维上,兴奋是可以双向的进行传导的,而且在这个传导的过程当中,主要是通过离子通道的打开和关闭等等来完成,他不会出现 信号的递减,所以一是双向的,第二信号应该是属于不递减的。 但是我们又要给大家提一下,在实际一个反射活动的过程当中,由于接受到刺激一般总来自于感受器,所以这样的话,沿着反射弧的路径,兴奋的传导应该是单向的,而不 是属于双向的。接下来我们看第二个问题,那就是兴奋在神经细胞之间怎么进行传递? 呃,由于我们说在神经纤维上传导的是一种局部的电流,现在如果是神经细胞和神经细胞中间就给断开了,那就相当于高速公路遇到了这一边河流。那么在这种情况之下,是怎么来把信号再给传送过去的? 这个时候就要借助化学信号了,所以我们在这边看一下,当兴奋传过来,首先将会引起这边突出小体发生一个改变,这个改变最主要体现在他的 这个突出前膜会兴奋,那么如果他发生了相应的兴奋,就会导致突出效泡发生移动, 移动到突出前模,然后这样的话,以包图的方式把地址是放到了突出间隙里边,而到了突出间隙里边之后,跟突出后模的相关的设计结合,引起后边兴奋或者是意志。 假如是兴奋性的低脂引起后膜的兴奋,如果是抑制性的低脂引起后膜应该是产生相应的抑制, 那么当这个相应的信号作用完了之后,他会失火。所有的信息分子当发挥完信号传递的任务之后,那么一般都应该要通过灭火的路径使他失去相应的作用的。那么在这边的灭火主要是什么方式呢? 可能是酶的作用,也可能是被回收,重新回收到突出前的这个突出小体里边去重新再来发挥相应的 作用。那么很多的药物的作用实际上就是作用在这个突出的这个相应的位置。所以大家可以想一下,如果某一种类型的物质是作用于这一边的话,它可以作用于什么呢? 他可以影响到这边地址的释放,如果影响到地址释放,那当然就影响到了相应的信号,对吧? 他可以影响到兽体,因为兽体是跟地质进行结合的,我假如有一种物质是跟这个兽体结合了,他本身就有地质的效应,那后续的效应是加强。如果他本身不具有地质的效应,那就构成了一个竞争的关系,那么信号的作用就会相应减弱, 他还可能影响到地质的回收或者是灭火,对吧?所以这样的话,我们就有了一个思考问题的相应的点, 前面我们说了,在兴奋神经纤维上传导的时候是双向的呃,而且他是不递减,速度也应该是比较快的,而在这边的话,他就变成这个单向的过程, 而且由于他要把电信号转化为一种化学物质的信号,然后再进行转化为电信号,所以这样的话他会有个时间的耽搁,因此这样的话,他将表现为单向和速度相对是比较慢的这样一个基本的特点。 那么接下来我们看下边一个题目,这个题目说下图是反射弧的一个局部的结构示意图,刺激的是 a 点,然后 a 点为两个连接线的一个中点的位置,然后检测各个定位的变化。下列说法错误的是好,首先我们看 哎,电表一是不偏转,然后电表二可能偏转两次。首先这个中拉音是中间的部分,他到达这个一这个部分的两侧,这个电极的连接的位置应该是距离是相等的,所以两者同时兴奋这个指针是不发不能偏转吧。 信号沿着这个突出传到第二个这边过来,那么先是左边的这一边,兴奋指针一次偏转,然后传过来到右边的这个电机的位置,再发生一次偏转,两次方向相反的偏转,所以 a 应该是属于对的。 谁呀?兴奋有 c 传到 e, c 到 e 的过程当中发生了电信号化学性好和电信号的一个转变,那么如果是跨过了相应的突出,就意味着有化学 信号的一个过渡,所以这有一个突出在这边,所以也应该是属于正确。 若深处检测不到定位的改变,可能是有突出前释放了抑制性的地质,那么假如前面是一个抑制性的地质,释放后膜就不会兴奋,所以在这种情况之下就检测不到相关的定位的改变,也应该是合理吧。 第四,若检测到 bd 有膜定位的这种变化,说明兴奋在同一神经元上是属于双向的, 优异的刺激是在 a 的这个位置,所以沿着这个信号过来,到 b 到 c 到 d 到 e 应该是一个方向进行传递,而不应该是属于双向的。呃,所以在 b 和 d 两个地方 是能够检测到膜定位的改变,那是因为他有个时间差在里边,所以这样的话我们说错误的应该是属于时的。好,那我们在这一刻当中主要就给大家分析到这,同学们,再见。
2141淼森 44:27
44:27 11:40查看AI文稿AI文稿
11:40查看AI文稿AI文稿同学们好,这节课我们继续学习高中生物文态与调节的第二章,神经调节。本节课的核心内容是神经冲动的产生与传导。三、 第一,神经元的连接方式。上一节课呢,我们已经给同学们介绍了呃,神经元的一些结构,大家知道神经元啊,包括包体、竹突、数突、神经末梢等。 那么神经元与神经元之间是怎么样相连的呢?他有两种连接方式,第一种叫逐突包体连接, 也就是说啊,这个神经元的竹突的神经末梢,他连接在下一个神经元的包体 上啊,在这里啊,这叫逐突包体连接。第二种连接方式呢叫逐突竖突连接, 就是说这个竹兔的神经末梢,他连接在另外一个神经元的数图之上啊,这个神经元连接的方式呢,大约就是这两种,他们都会形成一个结构,叫做突出。 那么我们来看一下这个突出的结构啊,你看,这就是相连接的这个点,他有什么样的结构呢 啊?放大以后就是这个样子,我们一起看一看啊。首先这个像葫芦一样形状的这个结构叫突出小体,突出小体当中 最核心的就是这个突出小炮,突出小炮当中啊,有一些神经地质存在, 这是一些化学物质,能够起到不同的作用,然后这边呢有线粒体为神经递质的释放提供能量, 然后这个神经地质就装在这个突出小炮之中啊,他有起兴奋作用的,也有起抑制作用的, 然后下方这个膜就是突出前膜,那么神经地质的释放就是在前膜之上以包吐的形式被排放出来的, 中间的这个东西呢叫突出间隙,突出间隙实际上他 本质就是组织液,这里边呢含有很多的啊,一些无机烟离子啊,水啊,蛋白质之类的东西。 然后下方的这个膜呢,就是突出后膜,然后突出前膜,突出间隙,再加上突出后膜,三者就构成了我们所说的突出, 也就说两个神经元之间一旦接触在一起的话,就有这样的一个结构啊, 然后这里面突出后膜,大家要明白,有一个非常重要的结构叫瘦体, 它是属于糖蛋白的一类啊,它可以识别神经低质的种类。然后呢,识别了以后,突出后膜 就会发生不同的反应,有的呢维持外正内负的静息状态,而有的呢就是变成了外负内正的兴奋状态。 那么我们探究一下兴奋在神经元之间的传递过程。首先第一个问题,兴奋在神经元之间传递依赖的结构是什么?这当然就是突出的结构, 该结构有哪几部分组成三部分,突出前模,突出间隙,突出后模。 第二问,兴奋在神经元之间传递依赖哪一种化学物质 啊?依赖的化学物质呢,就是神经递质,他有的起兴奋作用啊, 有的企业抑制作用。那么发生了哪些信号的转换呢?这个信号的转换,首先是电信号到化学信号,再到电信号的转换,在突出钱膜之上 发生的是电信号到化学信号,也就是说逐突传导过来的这个生物的电流,引发了突出小体当中神经递质的释放,这个过程就是电信号到化学信号的转换。 而在突出后膜之中呢,就是化学信号重于突出后膜当中的受体,以后啊受体他识别了这个化学信号,引起突出后膜 有些电流的产生啊,这个就是电信号,所以在整个突出之中,就是电信号到化学信号再到电信号。 那么第三,兴奋在神经元之间单向传递的原因是什么? 你看我们上节课讲神经纤维啊,兴奋在神经纤维上的传导是双向的, 你在神经纤维的中间进行一次刺激,这个电流朝两端跑, 但是兴奋在神经元之间却只能单向传递,其根本原因啊,就是神经递质只能由突出前模式放啊,到达突出间隙,作用 于突出后魔。所以我们说兴奋在神经元之间是单向传递的,这也就是 为什么在反射弧之中,这个信号的传导只能有感受器到传入神经,到神经中枢,到传出神经再到相应器的变化了。是因为在反射弧当中有很多的这个突出的结构。 那么神经地质的种类和功能有哪些呢? 种类有兴奋性的地质,比方说以显胆碱,他可以使突出后膜形成外腹内症的动作 点位,也就是说,人在工作、学习、运动的时候,他这个神经地质的释放就是以兴奋型的神经地质为主了。 第二类呢,是抑制性的神经递质,如干氨酸,它可以使后膜维持,甚至加强外症内负的静息定位。这样以来呢,人在休息睡眠的时候,这可能就是以抑制性的神经递质为主了。 那么神经递质的释放方式,不论是大份子的还是小份子的神经递质,他的出去方式都是包吐。 第三,去向,神经地质被突出后磨 的受体识别了以后啊,他发挥到了一定的作用,然后呢,就迅速被酶类啊给降解了,或者说有一部分呢,被回收进细胞了,以免持续发挥作用。 那么大家可以想象一下啊,这个神经递质如果持续发挥作用会导致什么?那我们的肌肉就得不到休息了啊,会不断的颤抖。 第四,药物影响神经递质机制的分析。我们主要介绍三类啊, 第一类啊,某些药物,如见毒,就是猎人射箭啊,在那个箭尖上抹的毒药啊,他可以与突出后膜上的受体结合,导致受体无法识别, 别神经抵制,这样以来呢,就会导致猎物被射中以后呢,这个肌肉松弛,趴在那不能动。 第二,某些药物,如部分的农药,它可以抑制分解神经地质酶的活性,使神经地质持续作用于突出后膜上的受体 啊,这样以来呢,就导致突出后膜持续的兴奋,导致肌肉僵直震颤啊,比方说有些啊,这个农药中毒的案例就是这个样子啊。 第三类,药物止痛的原理。这个止痛药,它的作用原理是什么呢?药物与神经地质,它可以争夺突出后膜上的 特性受体,但是这个争夺啊,他不是说永久性的争夺啊,只是暂时性的争夺,他可以阻碍兴奋的传递药物。呃,这样以来,神经地质的合成和释放就被干扰了, 神经地质的合成与释放一旦被干扰,那么人就感觉不到疼痛了。 那么第五,兴奋剂和毒品的作用原理。我们以科卡因为例来说明, 这个人的神经当中有一类神经地质叫多巴胺,多巴胺可以纵于突出后膜上的多巴胺受体,然后引发啊一定的作用,因为多巴胺呢,他可以使电呃,使这个大脑啊产生愉悦感, 多巴胺一释放,人就很快乐。而正常情况下呢,这个多巴胺应该是很珍贵的啊,他会被呃突出间隙当中的多巴胺转运蛋白给他回收了。 而可卡因呢,这个毒品可卡因,它会与突出间隙中的多巴胺转运蛋白结合, 这样以来呢,就使多巴胺转运蛋白失去了回收多巴胺的功能。那么这个多巴胺在突出间隙之中持续的纵于多巴胺受体,人就会不断的感觉到快乐。但是这个后果是什么呢? 多巴胺在突出间隙中持续发挥作用,导致突出后膜多巴胺受体减少啊啊,多巴胺受体 也受不了那么多的多巴胺,所以到了最后,多巴胺受体已经减少了,为了追求愉悦感,吸毒者会继续吸食科卡因啊,来维持这种快乐,结果就造成了恶性的循环 啊,他只要不吸这个东西,他就会非常的抑郁难受啊,不容易快乐了啊,所以这毒品呢,对人的危害是非常之大的。 好,我们这个本节课呢,就上到这里,那么下节课呢,将带同学们继续学习神经冲动的产生与传导,我们将会以典型立体的形式帮助大家巩固这一部分的内容。好,同学们再见啊。
600周浩老师 00:48查看AI文稿AI文稿
00:48查看AI文稿AI文稿很多学生不理解,为什么兴奋在神经纤维上是双向传导的,于是呢,我利用西沃白板就设计了一块可以移动的局部电流。蓝色图案,部分表示局部电流,通过拖动这一块 图形,然后向两边移动,表示兴奋,可以向两边进行传导。 这一个呢,是突出的结构,突出的结构也是比较复杂的。 通过这样的方式呢,可以简化学习的难度。而且让学生到平板上面进行拖动,可以提高学生学习的兴趣。学生自己有平板,也可以进行相应的操作。
177亲亲紫荆 00:21查看AI文稿AI文稿
00:21查看AI文稿AI文稿神经冲动传到突出前膜,促使钙离子内瘤引起突出小泡,与前膜融合并释放乙酰胆碱, 乙仙胆碱穿过突出间隙,作用于突出后膜的乙仙胆碱兽体,从而引起突出后膜,产生兴奋神经冲动就传到了下一个神经元。
603唯美生物 15:00查看AI文稿AI文稿
15:00查看AI文稿AI文稿各位同学大家好,我是来自北京大学附属中学的高成老师, 今天我们一起来学习高二生物学选择性必修一第二章第三节神经冲动的产生和传导的第一部分跟随科学家的脚步,破解神经冲动产生的具体机制。 通过之前课程的学习,我们已经知道神经调节的基本方式是反射,反射的结构基础是反射弧 反射。在短跑赛场上表现为发令枪一响,运动员们就会像离弦的箭一样冲出起跑线。运动员从听到发令枪响到做出起跑反应这样一种条件。反射感受器是分布在耳朵 的听神经末梢,通过听神经将信号传入神经中枢大脑,在大脑做出反应后,指力经过传出神经传导至腿部肌肉,也就是效应器完成起跑这一反应。 在这一过程中,兴奋在感受器如何产生,又是如何在神经中传导的呢?短跑比赛规则规定,发令枪响后零点一秒内起跑为抢跑。这一规定的生物学依据 又是什么?这些是我们这一节需要重点讨论的内容。对这一问题的探索肇事于意大利医生和生理学家加尔瓦尼。在一七八六年的一个偶然发现。加尔瓦尼发现挂在铁炸的铜 鸿沟上的蛙腿在风的吹动下左右摇晃,蛙腿一碰到铁炸了,就可观察到肌肉明显的收缩。 加瓦尼认为这种收缩是肌肉内部流出来并沿着神经到达肌肉表面的电流刺激引起的,即动物的组织可以产生生物电。 加瓦尼的观点在科学界引发了争论,福特等科学家认为加瓦尼的发现很有可能是铜铁两种金属的电位差引起的,而不是所谓的生物电。为此,加瓦尼和他的后记者设计了无金属收缩实验。 在挖坐骨神经肥肠肌标本中阶段挖的坐骨神经可以导致挖肥肠肌收缩。在这一过程中 没有设计任何金属,说明生物电确实存在。随后,电流剂于一八二零年应用于生物电研究。科学家在挖神经外侧连接两个电极, 随后刺激蛙神经一侧,并在刺激的同时记录电流表的电流大小和方向。在刺激刚开始时, 神经上电极所在的 a 点和 b 点均没有兴奋,故电流表不显示电流,说明 a 点和 b 点均没有兴奋,没有电位差异。当兴奋 传导至 a 点时, b 点所在位置还没有兴奋,可见电流表出现明显偏转,电流从 b 点流向 a 点,说明 明 a 点比 b 点的电位低。当兴奋传导至 b 点时, a 点所在位置已经由兴奋回复到静息状态,此时电流表出现明显偏转,电流从 a 点流向 b 点,说明 b 点比 a 点的电位低。当兴奋 传导至 b 点右侧时,兴奋已经传导过 a 点和 b 点,此时 ab 两点均为静息状态,电流表不显示电流,没有电位差异。 该项实验证明,兴奋在神经上以电信号传导神经,兴奋发生位置电位低于静息位置。那么,为什么神经兴奋发生 位置电位低于静息位置呢?在发生兴奋的位置是否存在跨生物膜的电荷转移呢?依据刚才的实验,科学家提出了生物电发生的魔学说, 因为生物膜具有选择透过性,神经兴奋的发生可能是生物膜调节假离子或者其他离子的透过性,进而调节细胞膜两侧电位差引起的。要回答这个问题是用蛙的神经是不可以的。 在学习神经调节的结构基础时,我们已经知道神经元轴突呈纤维状,外表大都有一层水翘结构,构成了神经纤维。许多神经纤维集结成数,外面包有一层包膜构成神经。因此,如 如果我们要观察神经兴奋是否有跨生物膜的电荷转移,需要测量轴突所在细胞膜两侧的电位差,这需要将一个电极插入轴突内部,要求电极的直径非常细,且不能损伤细胞。在当时, 这是一个难以逾越的技术难题。一九三零年,英国解剖学家杨发现一种软体生物枪乌贼的神经中单根轴突的直径异常粗大,是研究电声里的优秀生物材料。同时, 微电极的长足发展使得微电极直接插入神经纤维内成为可能。这幅图展示的就是插入枪乌贼轴突的微电极。一九 三九年,赫旭离和霍奇金使用微电极技术和细胞内记录的方法,研究枪乌贼神经细胞轴突膜两侧的电位变化。他们将枪乌贼的神经元轴突进入成有生理盐水的水槽,将其中一个电极刺入细胞膜,而 另一个电机留在细胞膜外,并将两个电机连通,监测电位变化和距离。和霍希金使用的微电机直径很细, 而且中空内部充满了生理盐水,在维持神经元轴突原有生物活性的前提下,能够有很好的导电性。 实验发现,在要插入枪乌贼轴突的微电极刺穿轴突细胞膜前,两个轴突之间没有电尾差异。在该电极刺穿细胞膜 后,两个电极之间出现了四十五个毫伏的电位差异,且强乌贼轴突细胞膜内电位低于强乌贼轴突细胞膜外电位确认了内负外症的近期电位。 在此基础之上,赫距离和霍希金开始探究兴奋在轴突上的产生机制。神经在 在未受刺激时,细胞内液的假离子浓度远高于细胞外液,而细胞内液的钠离子浓度远低于细胞外液。 我们知道细胞膜具有选择透过性,此时细胞膜内外的钠离子和假离子不能在膜内外自由往来,细胞膜两侧电位表现为内附外症,这种电位称为 静息电位。同学们回忆一下我们刚刚提到的魔学说,细胞内液钾离子浓度远远的低于细胞外液轴突。在静息状态下,细胞膜内电位低于膜外。若静息时, 细胞膜只对假离子有通透性,由于带正电核的假离子顺浓度差向细胞外扩散,相应的负电核仍留在细胞内, 这样细胞膜两侧形成了外正内部的静息电位。神经受到刺激兴奋时,细胞膜受到刺激,瞬间失去了对假离子的选择性渗透,导致对所有离子通透,两侧的电位差瞬间消失而形成 兴奋。如果魔学说成立,贺旭离和霍奇金在刺激枪乌贼轴突后,应该观察到怎样的电位变化呢?魔学说认为,神经静息状态的维持和兴奋的产生主要依靠细胞膜对假离子的调控给予刺激时, 魔两侧的电位差瞬间消失,形成兴奋。因此,如果魔学说成立,在枪乌贼轴突受到刺激时,魔两侧的电位差应从四十五毫伏变为零。左图是 特叙利和霍奇金记录的给予刺激后枪乌贼轴突的电位变化。那么记录的枪乌贼动作电位符合魔学说吗?我们可以观察到,轴突 兴奋的时候,电位变化的幅度大大超过了零电位,而到达了正三十到五十毫伏。 那么,轴图兴奋时是如何形成正电位的呢?按照魔学说的观点,如果仅仅有假离子的外流, 不会形成细胞膜内正电位和距离。和霍去金进一步的研究在一九五二年发表的论文中指明了动作电位形成的原理。 在这篇论文中,贺旭离和霍奇金指出,第一,兴奋的产生,一, 依赖纳离子的内流和假离子的外流。第二,动作电位发生时,细胞膜内电位迅速升高,由纳离子内流决定。 第三,纳离子由细胞外向细胞内的大量内流决定细胞膜内的电位变化,并导致膜内电位为正。 第四,动作电位恢复为静息电位时,假离子外流具有关键作用。在研究中,他们突破性的发现了纳离子在兴奋形成中的关键作用,为动作电位形成时细胞膜内正电位的形成找到了合理的理论依据。 而魔学说中具有关键作用的假离子则在动作电位形成后,通过假离子大量外流,使得刺激位置恢复为静息电位。纳离子和假离子两种离子共同参与了神经刺激部位兴奋的形成和 静息电位的恢复。一九六三年霍齐金和贺旭离在众多电位发生机制上的卓越工作, 与另一位神经生理学家埃克尔斯共同获得了诺贝尔生理学与医学奖。生物电现象和动作电位发生机智的发现很大程度上依赖于研究手段的不断深入和更新。 电流器可以帮助我们直接观察和记录电活动。微电极技术和后续电压。前和膜片衔技术的发展使我们能够在维持神经轴突活性的同时,将电极插入细胞膜内, 在细胞水平帮助我们认知生物电现象的本质。腔污贼粗大的神经元轴突发现则使我们具有理想的生物实验材料。技术 不断迭代和发展,帮助我们不断深化对生物学的本质认知。距离破解生物的奥秘越来越近,科学的发展最高境界是使用数学语言来表达自然的本质运行规律,生物学距此还有很远的距离。 霍奇金和贺旭梨通过研究动作定位发生机制而形成的霍奇金、贺旭梨方程,是生物学研究历史上少有的能够使用数学语言表达的生物运行规律,有兴趣的同学可自行深入学习。 在生物学研究历史上,霍奇金和霍旭梨因霍奇金、霍旭梨方程与将数学首次引入生物学研究的孟德尔具有相同的历史地位。神经纤维未受到刺激,细胞膜两 两侧电位表现为内附外症的静息电位。当神经纤维某一部位受到刺激时,细胞膜一部分纳离子通道开放,对细胞外纳离子的通透性增加 纳离子内流,细胞膜内电位开始升高。如果刺激足够强烈,当细胞内电位从静息电位升高到一定程度时, 细胞膜上的大量纳离子通道开放,从而使得纳离子大量内流推动细胞膜内电位迅速升高,使得 细胞膜两侧定位由静息定位的内负外正转变为内正外负,从而形成动作定位。诱导细胞膜大量纳离子通道开放的定位称为预定位。只有刺激诱导 纳离子内流,使细胞膜内电位到达预电位,才能够诱导动作电位形成,使得神经纤维刺激部位兴奋。 动作电位形成后,神经纤维刺激部位需要及时恢复为静息电位,以便及时感知后续刺激。 此时细胞膜上的纳离子通道关闭。由于此时细胞膜内假离子浓度远远高于细胞外,细胞膜上的假离子通道开放,大量假离子外流,从而使细胞膜内电位快速下降,从内正外负的动作电位重新恢复为内 负外症的静息定位。在恢复为静息定位后,神经纤维刺激部位需要产生动作定位过程中,大量内流的钠离子通过主动 运输的形式运输到细胞外,将由动作电位恢复为近期电位过程中,大量外流的假离子通过主动运输的形式运输到细胞内,从而恢复原有的细胞内外纳离子和假离子的离子浓度分布,以便在之后刺激时能够继续形成动作电位。 现在我们已经知道,神经处于静息电位时,神经细胞膜外的钠离子浓度高于细胞膜内 神经细胞膜外的钾离子浓度低于细胞膜内。清晰定位下细胞膜内外的钠、钾离子浓度差如何维持呢? 动作定位发生后,大量钠离子自细胞膜外内流至细胞膜内,大量假离子自细胞膜内外。
161淼森 06:44查看AI文稿AI文稿
06:44查看AI文稿AI文稿同学们好,这节课我们继续学习高中生物稳态与调节的第二章神经调节。本节的核心内容是神经冲动的产生与传导。二、 经过上节课的学习,我们知道在神经元的内部,假离子的浓度是非常高的, 而在神经元的外部呢,纳离子浓度是非常高的,这样首先 假离子外流会导致静息电位,也就是外正内负的形成, 这个过程呢,是从高浓度向低浓度,这是一个协助扩散,而一旦有了刺激以后呢,纳离子通道 就打开了,这样以来纳离子就会从细胞外向细胞内进行运输,导致动作电位的形成,也就是局部的外副内症。 这个过程呢,也是从高浓度向低浓度,需要载体,不需要消耗能量的,所以他也是协助扩散。 而到了最后,呃,有一种非常特殊的载体啊,叫纳甲泵,纳甲泵呢,在消耗能量的作用之下,再把出去的假离子给他运进来, 进来的纳离子呢,再给他排出去,这个过程就是一个主动运输了,然后神经元的内外通过这三个过程导致了进吸 定位,动作定位以及离子的平衡。 我们来具体的看看这个过程, 现在是惊喜典韦,大家可以看见红色的这个颗粒代表的就是假例子,他开始从内向外排出去了,这是一个 不需要消耗能量的协助扩散课程。 而当刺激产生以后呢,那粒子通道打开了, 这时候蓝色的颗粒,他代表的就是纳厘子,他不断的从外面向内边流动,也是协助扩散。 而随后有一种非常特殊的载体叫那家蹦, 他以主动运输的形式,将进来的纳离子再给他排出去,然后把假离子再给他运进来,一般来讲,排出三个纳离子,就会运进两个假离子,这就是主动运输, 然后这个离子的进出,导致了这个静息电位与动作电位,他会形成一个 这个动作电位的这样一个图。波形图,我们来仔细分析一下这个波形图啊。首先在 a 点,他是静息电位, a 点是静息电位,他的主要特点呢,是外正,而内负是由嗯,假离子外流引起的,这个时候呢,那离子通道是关闭的, 而随后到达必点以后,刺激产生了,刺激一产生,这个假离子照样外流, 而与之不同的是,纳离子的通道打开了,很多的纳离子迅速内流,纳离子一内流啊,这个外正内负的电位就变成了 局部的外负而内正,而随后由 b 点到 c 点呢,会有啊,这个假离子外流,但纳离子内流会不断的增多,这样以来,到达地点就是一个顶峰了,地点 那离子流失的已经很多了,这个时候呢,他的那离子通道就关闭了,动作定位外负内正啊,这个就达到一个极限了, 随后由地道溢,假离子仍然外流,而且流的更多了, 这个时候呢,那离子通道仍然是关闭的,那么假离子外流一多了,就会使原 先的这个外正内负的电位再一次形成。 而与此同时呢,纳离子和假离子的特殊载体叫纳甲泵,开始吸甲排纳,维持离子的平衡, 大约就是这么一个过程。那么 a 点呢,是外正内负,我们可以给他称之为极化现象啊,外正内负是极化,而从 b 点,一旦词迹以后,大量的纳离子内流就抵消了这个外正内负 这个定位,使之变成了外附而内正,那么这个现象呢,就叫趣极化啊,趣极化到达顶点之后,就开始 是恢复外正内服的进气垫位了,所以这个过程呢,就叫负极化。 由于大量的假离子外流,最后会导致这个静息电位啊,比之前还要强一些,就这个一点的静息电位比 ab 两点还要强一些, 这叫超级化现象。而随后从 e 到 f, 最主要的就是纳甲泵再进行起作用,维持啊这个离子的平衡。 好了,本节课就上到这里,那么下节课呢,将带同学们继续学习神经冲动的产生与传导。如果对本节课有任何的疑问,可以发表评论或者与我交流,我会尽快为大家解答好,再见!
586周浩老师 15:32查看AI文稿AI文稿
15:32查看AI文稿AI文稿同学们好,这节课我们继续典型立体的形式,帮助大家巩固高中生物稳态与调节的。第二章第三节神经冲动的产生与传导 那么题组四突出的结构第一题二法银环蛇毒 能与突出后膜上的以显胆碱受体牢固结合,有机灵农药能抑制以显胆碱纸酶的活性, 而一线胆碱纸霉的作用是清除与突出后膜上受体结合的一线胆碱。因此,二发银环蛇毒与有机灵农药中毒的症状分别是。那么通过我们刚才的提干,这个二发 银环蛇毒它既然能与以显胆碱受体牢固结合,那么以显胆碱就没有办法作用于突出后膜了。因此啊,二发银环蛇毒它不会使兴奋产生, 而有机灵农药呢,它降低了胰腺胆碱姊妹的活性,这样以来,胰腺胆碱姊妹就不能够把发挥作用的胰腺胆碱给清除了,所以它会使得我们的身体持续产生兴奋, 这样以来,对应的症状就是,不产生兴奋的话呢,那就是肌肉松弛,而持续产生兴奋呢,就是肌肉僵直。所以本题呢,应该选 a 答案,第二题 以下有关神经元及兴奋的叙述中,正确的是。 a 选项, 兴奋状态时,神经元细胞内外也有离子进出啊,这个静息状态时,静息状态时有没有离子进出呢?这是有的,它主要是假离子的外流,而兴奋状态呢,主要是纳离子的内流。 b 选项,静息状态下,神经细胞膜两侧的垫位为外负而内正, 这个静息状态下,这个电位啊,是外正而内负,外负内正呢,是 兴奋状态下的定位。 c 选项,神经地质与突出前膜上受体的结合具有特异性,这个神经地质结合的不是突出前膜啊,而是突出后膜。 d 神经递质一旦纵容于突出后膜,一定会使其兴奋啊,这是错的,产生兴奋或者是意志。所以本题呢,只有 a 选项是对的。 第三题,如图为突出的亚显微结构, m i 音分别表示两个神经元的局部结构,下列相关叙述正确的是。那么我们先对这个图像呢进行一个精细 的加工,我们看看各个部位分别是什么。首先,小一代表竹突的啊,这个末端小二是提供能量的线立体,小三是含有神经递质的突出小泡, 小四是突出前膜,小五是突出间隙。它的主要成分呢,是组织液,小六是突出后膜,上面含有识别神经地质的受体。所以 a 选项 一、二、三合成突出小体是神经,是神经元数图的末段啊,检验错了,应该是逐图的末段。 b, a 点兴奋时, a 点 膜内垫位为正,而 b 点膜内垫位为负。哎,这是对的,因为呢, a 点一旦兴奋,原先膜内的负垫位 就会转变成正点位,而此时必点呢,还是静息状态,他依然是外正而内负啊,所以他的模内呢,为负点位。 c 神经地质存在于二中啊,这显然错了,神经地质应该存在于三之中啊,二是线立体提供能量的,三是突出小泡内部含有神经地质五处的液体为组织液啊。当然,这句话呢,是对的。 b 选项,神经地质通过四的方式为自由扩散,那么神经地质通过突出前膜呢, 为包兔而不是自由扩散。所以本题呢,我们选啊 b 答案第四题, 神经地质分为兴奋性神经地质与抑制性神经地质两种,如以显胆碱可作为一种兴奋性神经地质,而去甲肾上腺素可作为一种抑制性神经地质。 那么下列说法正确的是,我们先分析一下啊,兴奋性帝制,他 主要的一个作用呢,就是使突出后膜形成外副内症的动作点位。而抑制性神经地质呢,它使得突出后膜维持 外政内副的近期电位。所以 a 选项, 二者由突出前摩进入突出间隙时,都需要借助载体的运输啊,这是错误的,因为神经地质的释放啊,不论大分子还是小分子啊,他一般来讲呢,都是包吐,所以说是不需要借助载体的啊。 b 二者都能够被突出后膜上的受体识别,体现了细胞间信息交流的功能啊,没有问题。 c 二者都能够长时间,重于突出后膜,使膜垫为长时间发生改变。那么错就错在长时间啊,呃 呃,兴奋性的神经帝制,他作用的时间呢,是非常短的啊,他使得膜后突出后膜的垫位呢,由外症内负变成了外负内症,这个时间是相当短的。 而静息电,而这个抑制性的神经递质啊,他不会使电位发生改变的啊,他一直使的电位呢,维持在外正内负的这样一个状态。 d 选项,二者重于突出后膜后细胞膜对假离子、纳离子的通透性都发生改变,产生动作定位啊,这显示错的。 兴奋性的神经抵制产生动作变味,他会使得纳离子通透性改变,迅速内流啊,而 抑制性的神经病,抑制性的神经递质啊,他不会产生动作电位的,他是维持近期电位的。当然,这个时候呢,假离子他的通透性可能改变,会使更多的假离子外流。所以本题呢,我们选 b 答案 第五题,氨基丁酸 gaba 作为哺乳动物中枢神经系统中广泛分布的神经递质,在控制疼痛方面的作用不容忽视,其机理如下图所示。 下列对此判断错误的是,那么既然提干当中说了氨基丁酸啊,他可以控制疼痛,所以我们预判断他一定起到的是抑制性的 作用。然后再分析图像呢,大家可以看见,氨基丁酸进入突出间隙以后啊,他可以与绿离子的通道进行结合,使很多的绿离子进入 啊,这个绿离子一旦进入啊,实际上就维持了外正内负的进气垫位啊。所以这个图像进一步使我们确信,氨基丁酸起到的就是抑制性的这样一个作用。所以 a 选项,当兴奋到达突出小体时,突出前膜释放 gaba, 该过程依赖突出前膜的流动性啊。对的,因为包吐呢,他就是建立在 包膜流动性的基础之上的。 b, 突出前膜释放 gaba 的过程,说明某些小分子物质可以通过包吐的方式分泌出细胞啊,这个也没有问题, cgaba 受体实际上也是横跨突出后膜的绿离子通道,能与 gaba 特性结合啊,这个也没有问题。 d jba 与受体集合后,会导致绿离子内流,竟然导致突出后膜产生动作电位,哎,这个错了,应该是维持或者是加强了静息电位。 所以第五题呢,我们选择的是递答案。 第六题, 去甲肾上腺素 n e 与多巴胺 d e 结构相近,是一种兴奋性的神经递质,小剂量可以升高血压。 去甲肾上腺素也是一种激素,由肾上腺随致合成和分泌。下图表示肾上腺突出中化学变化及兴奋的传递过程。也就是说啊,当这个 肾上腺突出啊,就神经细胞纵于肾上腺啊这样一个受体。以后呢,大家可以看到,突出前膜之内,由络氨酸进步转化成多巴,然后再添加化学集团转变 多巴胺,随后有突出小泡,内部多巴胺还有可能演化成去甲肾上腺素。 然后呢,它作用于去甲肾上腺素受体,引发身体的一个变化。而随后 这个释放出来的恩义啊,他有可能被回收啊,通过这个一定的载体,他回收了,回收以后呢,他还有可能再一次进入到这个突出小炮内,等待下一次的释放。所以第一小问, 恩义的释放过程,他是一定要消耗 adp 的,因为这个过程呢,他是一个包兔的作用,恩义发挥作用后,回收 突出前膜的神经元,其意义是它可以避免恩义持续发挥作用啊,因为人体呢,也不能承受住持续的兴奋。 第二,恩义可用于治疗急性心肌梗塞等引起的低血压,通过于去甲肾上腺素受体结合增强 心肌血管的什么来升高血压。这毫无疑问应该是收缩,因为我们的血管一旦收缩,那么血压呢,就会升高。 在人体内,该物质可由交感神经元合成,可分泌,判断的依据是去甲肾上腺素会使血压升高,我们 交感神经,它主导的是人体的兴奋状态,它会使得血管收缩,疑使血压升高,而反过来呢,这个负交感神经啊,它会使得我们的血管扩张,然后来降低血压。 第三,在有效的抗菌药物的治疗下,临床上常通过联合使用多巴胺和去甲肾上腺素来降低 血清乳酸降钙素原含量,提高中心静脉压,从而治疗有微生物或者是毒素引起的感染性休克及其并发症。某科院人员以小暑 为实验材料,与通过实验比较多巴胺与去甲肾上腺素治疗感染性休克及其并发症的效果,那么该实验需设置极阻实验, 要设足四组实验,首先,第一组要空白对照,不加任何的这个多巴胺,或者是去加肾上腺素,用生理盐水来进行实验。第二组只加多巴胺,第三组只加等量的肾上腺素。 第四组既加多巴胺又加这个去甲肾上腺素,就是使他联合使用吗? 而这个阴变量呢,这个自变量就是多巴胺,呃,去甲肾上腺素有没有添加以及他 添加的方式?而因变量呢,就是我们检测的是小鼠体内的这个什么血清乳酸降钙素原含量, 还有小鼠内的中心静脉压的变化。因为这个知识呢,在我们的提干当中已经有了提示了, 所以你看我们生物学啊,他的考试啊,有点意思啊,就是答案往往是在题目之中的,大家一定要仔细的去分析题目,你可以从中找得到答案。 那么第二个,若联合使用效果优于单独使用,则联合组血清乳酸的含量怎么样?那么题目已经说了,这个激素可以降低血 血清乳酸啊,什么降钙素元的含量,所以说他如果效果很优的话呢,那么这个血清乳酸的含量就会明显低于单独食用族。 好,本节课就上到这里,那么下节课呢,我们将继续带同学们进行神经冲动产生于传导的点力经验。如果对本节课有任何的疑问,可以发表评论或者与我交流,我会尽快为大家解答好,再见!
181周浩老师 04:48查看AI文稿AI文稿
04:48查看AI文稿AI文稿动作垫位是可兴奋细胞,例如神经细胞在静心垫位基础上接受有效刺激后产生的一个迅速、可逆、可向远处传播的磨垫位波动。 动作定位包括快速去即化和快速负极化的风定位和其后缓慢波动的后定位。 风电位是细胞兴奋的标志。动作电位中风电位的产生与细胞膜对蜡假离子通透性的改变密切相关。 在静息电位基础上,当细胞受到有效刺激后,细胞膜首先对纳离子的电导及通透性增大,带镇定赫的纳离子在其浓度差和定位差的电化学驱动力推动下 快速流入包内,并引起膜的聚集化。当聚集化达风时时,膜对纳离子通透性降低并消失, 假电脑及膜对假离子通透性增加,于是那内流停止,假离子在其浓度差和电位差决定的电化学驱动力推动下快速流向细胞外,从而引起膜的负极化。 细胞膜对纳甲离子的通透性的改变与膜上电压门控纳通道和电压门控假通道所处的功能状态有关。 电压门控那通道具有激活门 m 和失活门 h 两个闸门。当细胞处于安静状态时,那通道激活门关闭,失活门开放。 通道处于静息状态,磨电位保持在静息电位水平。 当细胞受到有效刺激磨去即化,达到预定位时,大量的电压门控那通道由静吸肽转化为激活肽,这时那通道的激活门快速打开,吃活门则缓慢关闭。 在激活门已打开、失活门还未关闭的一至二毫秒内,膜对纳离子通透性突然增大,纳离子快速进入包内,使膜迅速发生雀迹化, 激活后的那通道随时机延续,失活门逐渐关闭。这时尽管激活门已打开,但那通道也不倒通, 称之为失火状态。电压门控。假通道只有一个激活门,恩门 细胞安静时其关闭为静息状态,去即化时可以激活,但开放较纳通道激活门慢的多,多数在纳通道失活后才开放。为延迟激活 电压门控,甲通道开放后,膜对甲离子通透性增大,引起的甲离子外流促成了膜的快速负极化。动作电位产生后,可以在局部电流的刺激下在同一细胞上进行不衰减传导。 在无损神经纤维及大多数细胞,动作定位的传导是顺序式传导。在动作定位发生的兴奋 膜两侧呈现暂时的外腹内正反极化状态,而其相邻的内心分区则仍处于外正内负的极化状态。 由于电位差的缘故,在细胞内将会出现由兴奋区流向临近未兴奋区的局部电流。相反,在细胞外局部电流的方向是由临近未兴奋区流向兴奋区。 在局部电流刺激下,临近卫星分区发生趣迹化,当达到预电位时,即可在该部位触发新的动作电位, 而原来的兴奋区则进入负极化过程。其后新的兴奋区又与其前方的安静区继续形成局部电流,并使之发生兴奋, 最终是动作垫位由近极远的传播开来。在有损神经纤维被水撬包裹的区域,缺乏电压门控那通道。裸露的狼飞结处电压门控那通道密度较高, 因此局部电流只能在兴奋区的浪费节与临近安静区的浪费节之间发生, 这种动作定位从一个狼飞碟跨越节尖区跳跃到下一个狼飞碟的传导方式称为跳跃式传导。
470生物侯老师 45:30
45:30 09:43查看AI文稿AI文稿
09:43查看AI文稿AI文稿同学们好,这节课我们继续典型立体的形式,帮助大家巩固高中生物文态与调节的。第二章第三节神经冲动的产生与传导我们看题组五、电表偏转问题。 第一题如图,在一根神经纤维的 ab 之间接一个电流剂,然后在神经纤维上的某点给予刺激电流剂的指针,如图所示。下列说法正确的是 我们假设 c 是 ab 的终点,那么如果你单纯的刺激 c 点,会使得 ab 两处同时有原先的正点位变成负点位,这样以来呢, 电尾叉仍然是不存在的,这个电表的指针是不偏转的。而如果在 c 的左侧进行刺激, 会首先导致 a 处由正电位变成负电位,而此时 b 处仍然是正电位,那么这个电表呢,会先向左偏转,而随后 a 处恢复成正电位, b 处呢,就变成了负电位了。 立柱变成负电位以后呢,这个电表就是向右偏转了。所以我们总结一下啊, 在 c 点的左侧刺激是先左后右两次方向相反的偏转,再 c 点的右侧刺激是先右后左, 也是两次方向相反的偏转。所以 a 选项他说若在 ab 两点之间,此机电流记指针不会如图所示,这显然是错误的。 b 若在 ab 两点之间刺激,电流剂指针一定有一次吐湿的现象,这个也是错的,因为如果在 c 点上进行刺激的话,电流剂的指针是不动的。 c 选项若在 a 点的左侧吃鸡兴奋传导到 a 点。十、 电流剂指针如图所示,那么在 a 点的左侧刺激他会使得 a 点率先变成负点位。这个电流剂指针是向左偏转的,不是向。 而 d 选项若在必点的右侧刺激兴奋传导到必点时,电流记者指针如图所示啊,这就对了,所以本题呢,选 d 答案 第二题,下图为娃后直坐骨神经的示意图,在电极 a 的左侧给予适当的刺激。下列说法错误的是,那么在 a 的左侧刺激的话,这个电表呢,最终是两次方向相反的片转, 所以 a 选项对了。 b 蛋白酶会影响受刺激后动作电位的产生,这是对的。为什么呢?因为动作电 为的产生啊,需要纳离子持续内流,而纳离子持续内流呢,他需要借助膜上的载体蛋白, 所以我们用蛋白酶来处理神经纤维的话呢,他有可能把细胞膜上的蛋白质载体给他溶解了,这样以来呢,就会影响这个电位的产生。 c 刺激产生的电信号在 a、 b 之间的传递方向为从 a 到 b 啊,这个没有问题。 d 选项,兴奋在神经纤维上的传导方向与魔内侧电流方向相反啊,这是错的,应该是相同的,与魔内相同,与魔外相反。 所以第二题我们选 d。 第三题如图是用甲乙两个电流表研究神经纤维及突出上兴奋产生及传导的示意图。下列有关叙述错误的是, 那么我们看 a 这个电表啊,他是一头插在外侧,而一头插在内侧的。我们知道,在静息状态下呢,是外正而内负,这样以来呢,电尾插产生,这个电表会持续的向左偏转。 而以这个电路表呢,它是架在两个神经元之间的,他们有一个突出的结构。而对于突出,我们知道了啊, 神经地质只能从突出前膜释放到达突出间隙,再纵于突出后膜,这个是不能反过来的,所以我们具体的看一看。 a 选下静息状态下,假指针偏转,而以指针不偏转啊,这个没有问题。 b 选项,刺激 a 处时,甲指针偏转一次,而乙指针偏转两次啊,这个也是对的, 因为你刺激 a 处,会使得这个甲这个外侧由正变负,而内侧由负变正,这样以来呢,他会向右偏转一次,然后又恢复啊原先 状态。而这个刺激 a 处以后呢,这个神经兴奋,他是从左向右传导的,会使得这个电表呢,出现两次方向相反的片转。 c 选项,刺激 b 处时,甲指针维持原状,乙指针片转一次啊,这是对的,因为你刺激 b 处,这个神经冲动是不可能从后膜传导至前膜的,所以甲这个电表是不动的。然后呢? 呃,刺激壁处以后,这个地方可能会由正变负,而这个地方依然是正,所以这个电表会向右片转一次。 c 清除 c 处的神经地质,在刺激 a 处时,假指针偏转一次,而乙指针不偏转。那么如果把神经地质去掉了,我们再刺激 a 的话呢? 嗯,这个对于假证电表呢,他确实是只片转一次,他会使得电位发生一个由外正内负变成外负内正的这样一个变化,而遗址真的实际上他也偏转一次。 为什么呢?因为遗址针他会导致 a 处的这个定位变化,然后 b 处呢,依然是正点位,他会向左偏转一次。所以第三题我们选 d。 答案。 第四题,图甲为某一神经纤维的示意图, 将一电流表的 ab 两极置于膜外啊,在 x 处给予适当的刺激,测得电位变化。如图以所示,下列说法正确的是,那么既然 ab 是位于膜外,实际上他同时为证 啊,这是没有电尾插的,取出电表是不动的。而你一旦在 x 处给予刺激的话呢,我们知道这个电表呢,会先左后右呈现两次方向相反的偏转, 那么他在图椅当中呢,就形成了一个山峰,一个山谷啊这样的图形。所以 a 选项未受刺激时,电流表侧得的为静息电位啊,这是错误的,因为未受刺激时他没有电位差呀。 怎么测静息电位呢?静息电位的测的一定是一个插在膜外,一个插在膜内才能检测的到。 b 选项,兴奋传导过程中, ab 之间魔内电流的方向为,由 b 到 a。 我们知道魔内电流与神经冲动的传导方向呢,是相同的,所以依然是从 a 到 b。 c 在图椅的 t 三时刻兴奋传导至 b 的电梯处,这个 t 三正好是波峰与波谷的这样一个交替的地方, 而在此时呢,刚好兴奋传导之 b 处,使得我们的电位发生变化。 d 选项 a、 b 两处同时有静息电位转变成动作电位,这显示错的 a 处先变成动作电位,所以第四题的是选 c 答案 好。同学们,这节课我们就上到这里,下节课我们继续进行神经冲动与 呃他的产生与传导的点力经验。如果对本节课有任何的疑问,可以发表评论或者与我交流,我会尽快为大家解答好,再见!
288周浩老师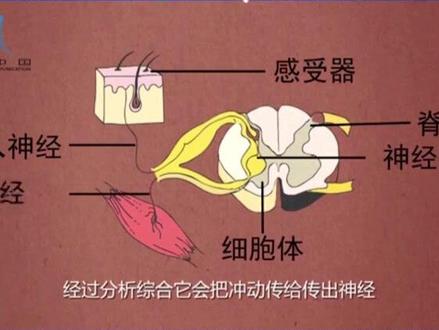 01:56查看AI文稿AI文稿
01:56查看AI文稿AI文稿当我们的手不小心碰到火的时候,会迅速不由自主的把手缩回来,这实际上是人体发生的一种缩手反射。 机体通过神经系统参与的对内外刺激都做出的规律性的反应,叫做反射。反射是神经系统活动的基本方式,它的结构基础是反射弧。 反射弧由感受器、传入神经、神经中枢、传出神经效应器五部分组成。以缩手反射为例, 他的感受器位于皮肤中,是传入神经末梢的部分。当他感受刺激后,会产生冲动,一种电信号并沿着传入神经传到神经中枢。神经中枢位于脊髓中,是神经元的细胞体集中的部分,经过分析综合,他会把冲动传给 传出神经,最后到达效应器。传出神经的末梢,加上他所支配的肌肉,引起肌肉的收缩,手就会缩回来,避免手被烫伤。 当信号到达脊髓后,也可以通过神经元传到大脑皮层,控制缩手反射活动。如果大脑下达指令不把手缩回来,就会把手烫伤,后果不堪设想。实际上, 偶然碰到火,等不到大脑的判断和思考,就会迅速的缩回来,这对我们机体起到了很好的保护作用。脊髓是低级的神经中枢,大脑皮层是高级的神经中枢。 缩手反射、吮吸反射、膝跳反射等都是人体生来就有的简单反射。神经中枢位于脊髓,人体还有经过长期生 活经验积累逐渐形成的复杂反射。要有大脑皮层的参与,反射活动的任何一部分结构损伤,反射活动就不能完成。如果缩手反射的结构中传出神经受到损伤,肌肉就不能完成收缩。
93邹晓科






猜你喜欢
- 9.0万燃尽